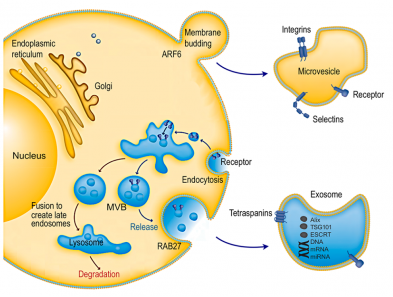
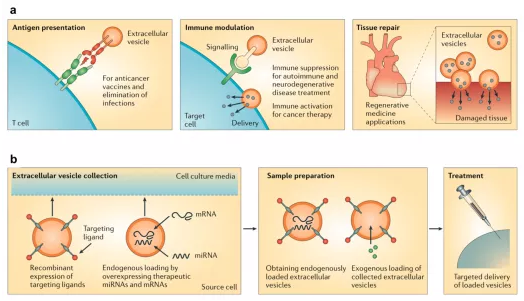
2.1SC-EVs对中枢神经系统损伤的修复治疗
血供受损和外力损伤是目前引起中枢神经系统损伤的常见原因,严重的中枢神经系统损伤常造成患者的死亡或残疾。由于神经元自行再生性极弱,中枢神经系统损伤后的功能障碍往往伴随患者终生,因此成为临床治疗中的一个需要突破的难题。神经元缺失、病灶血运受损及继发炎症损伤是中枢神经系统损伤后的基本病理改变,因此通过再生修复神经及血管并抑制炎症过度激活成为了中枢神经系统损伤一种治疗思路。然而利用外源性干细胞移植来修复神经损伤面临着血脑屏障阻挡、细胞存活率低及神经元分化数量低的主要难题。但同时研究也发现,尽管外周移植的干细胞常聚集于肺部,但仍能对中枢神经系统内的病灶起到促进神经和血管再生的修复作用,说明干细胞有可能通过分泌如SC-EVs等因子对损伤病灶产生治疗作用。
SC-EVs作为一种纳米级别的微小囊泡,可以有效穿透血脑屏障。利用SC-EVs促进颅内或脊髓内的中枢神经系统病灶的再生修复,是治疗中枢神经系统损伤领域的一个突破性研究方向。近年来的研究发现,独立的MSC-EVs对中枢神经系统损伤病灶中的内源神经元及血管内皮细胞具有促进再生作用。
这些研究分别在脑卒中、脑外伤以及脊髓损伤的大鼠体内发现,病灶中的神经元及血管内皮细胞数量在MSC-EVs治疗后显著增加,呈现出活跃的神经及血管再生现象。动物在MSC-EVs治疗后的神经功能试验表现及评分也表明,MSC-EVs能够改善中枢神经系统损伤引起的功能缺陷,具有明显的治疗作用。EVs中的miRNA是EVs发挥治疗作用的关键。
研究进一步发现,MSCs在缺血性损伤的脑组织中移植后,其分泌的MSC-EVs高表达miR-133b,后者对促进受损神经元轴突生长具有重要作用。已有研究也证实,MSC-EVs携带的miR-133b可通过激活下游ERK1/2、STAT3及CREB信号通路蛋白,在脊髓损伤病灶中对神经元的基因转录及细胞增殖功能具有促进作用,从而促进受损神经元轴突生长并缩小神经损伤病灶的体积。神经损伤后激活的炎症反应可能引起对病灶的二次打击。
有研究发现,脑组织内促炎因子IL-1β在脑外伤小鼠伤后6~12h内存在一个表达高峰。这种表达高峰所引起的急性炎症反应对早期脆弱的受损神经元会造成再次损伤。而MSC-EVs能明显抑制该因子在早期神经损伤中的过高表达,形成有利于受损神经元修复的病灶微环境。在治疗1个月内发现,MSC-EVs治疗组小鼠的记忆及认知功能明显优于空白对照治疗组。除了调节炎症因子,MSC-EVs对中枢系统中的多种炎症效应细胞也具有调节作用。
在脊髓损伤小鼠的治疗中发现,MSC-EVs能够降低病灶内星形胶质细胞的NFκBp65表达,并最终抑制星形胶质细胞在脊髓损伤后向具有神经毒性的A1表型活化。有研究发现MSC-EVs可以诱导上调小鼠脊髓损伤病灶内巨噬细胞M2亚型相关的ARG1及MRC1的mRNA表达,促使巨噬细胞向抑制免疫的M2亚型极化。
MSC-EVs通过对免疫平衡的调节改善了病灶微环境,有利于神经修复再生,在中枢神经系统损伤的治疗中具有重要作用。MSC-EVs除了具有促进神经血管再生及调节免疫的神经损伤修复作用外,对脑卒中治疗的最新研究发现,其含有的miR-134还可以抑制少突胶质细胞内caspase-8的表达。caspase-8是促进细胞凋亡的重要蛋白酶。少突胶质细胞是中枢神经髓鞘的重要组成部分,保证了神经电信号的正常传导。
MSC-EVs利用所含的miR-134抑制了因缺血而激活的少突胶质细胞凋亡,保持了病灶内该细胞数量的稳定。这对于维持中枢神经髓鞘完整性、修复脑内神经传导具有重要作用。由于利用胶原支架建立的三维培养环境能模拟自然组织结构,三维培养下的MSCs被发现对各组织损伤具有更强修复作用,对研究损伤修复治疗具有重要帮助。
在中枢神经系统损伤领域,最新研究通过对比脑外伤大鼠治疗后在水迷宫的逃脱时间,发现MSCs在三维培养下分泌的MSC-EVs比二维培养下分泌的MSC-EVs对大鼠空间学习能力恢复具有更好的效果。该项研究还发现前者含有的蛋白总量比后者含有的多一倍,这可能与三维培养下分泌的MSC-EVs具有更好疗效有关。在对中枢神经系统损伤治疗的研究中,MSC-EVs已被证实具有确切的促进神经血管再生、调节炎症环境、修复神经髓鞘等治疗作用。尽管相比MSCs移植治疗,天然的MSC-EVs没有表现出明显的疗效差异,但由于MSC-EVs具有相比MSCs更高的安全性和更强的可塑性,MSC-EVs在中枢神经系统损伤的治疗上更具有临床应用可行性。
2.2SC-EVs对阿尔茨海默病的治疗
阿尔茨海默病(Alzheimer's disease,AD)是一种常见的年龄相关神经退行性疾病,造成老年人的认知功能下降,影响自理能力及生活质量。迄今仍缺乏一种能够针对其病因或缓解其病理变化的有效药物,因此也是临床治疗的一大难题。尽管具体发病机制仍有待探索,但Aβ淀粉样蛋白在神经元胞外的聚集被认为是AD发病的其中一大原因。降解Aβ蛋白的堆积也是目前研究AD治疗的一个主要方向。MSCs能够分泌多种蛋白酶类,其中包括可以降解Aβ蛋白的脑啡肽酶。
近期研究发现,人脂肪来源的MSCs所分泌的脑啡肽酶被包含于MSC-EVs中。该研究进一步将MSC-EVs分离提取,并与过表达Aβ蛋白的AD病变神经元共培养后发现,MSC-EVs能够显著减少Aβ蛋白在神经元胞内胞外的堆积。首次证实了MSC-EVs对阿尔茨海默病具有缓解病理变化的治疗作用。Aβ蛋白过度堆积后,可对神经元诱导产生氧化应激等损伤,是引起AD患者认知缺陷慢性加重的重要原因之一。
最近研究发现,MSC-EVs能够降低体外AD海马体神经元中与活性氧相关的荧光信号表达,可以保护神经元免受Aβ蛋白诱导氧化损伤。另一项研究发现MSC-EVs降低了Aβ蛋白引起的诱导性一氧化氮合酶(inducible Nitric Oxide Synthase,iNOS)过高活性。由于iNOS的过高活性参与了Aβ蛋白诱导的神经元氧化损伤,MSC-EVs的这种抑制作用能够缓解病变神经元所受到的氧化损伤。该研究进一步发现,在AD转基因的小鼠模型体内,MSC-EVs治疗能够明显改善小鼠在AD病程中的认知缺陷恶化,证实了MSC-EVs在活体动物体内对AD具有治疗作用。
最新研究同时发现,MSC-EVs对Aβ蛋白相关损伤的神经元也具有保护与促进再生的作用。MSC-EVs通过抑制由Aβ蛋白引起的突触蛋白PSD-95减少,从而保持海马体神经元突触数量稳定并促进突触生长。由于海马体内神经元的突触密度与动物认知功能密切相关,MSCEVs该修复作用对治疗AD认知缺陷具有重要价值。另一项研究发现Aβ蛋白堆积能引起AD转基因小鼠脑内促凋亡因子p53、Bax及caspase-3的高表达,而降低抗凋亡因子Bcl-2表达。这种凋亡相关因子的失衡最终导致了神经元的大量凋亡。
MSC-EVs能降低促凋亡因子的高表达,并提高抗凋亡因子的低表达。在抑制神经元凋亡的同时,MSC-EVs还对病变损伤的神经元具有显著的促进轴突再生的作用,这与MSC-EVs在缺血或外伤等急性神经损伤中的治疗作用十分类似。此外,MSC-EVs对Aβ蛋白激活的小胶质细胞免疫应答也具有调节作用,能够改善AD脑组织中的神经元生存微环境。
研究发现MSC-EVs在AD转基因小鼠体内能够抑制小胶质细胞向促炎M1亚型极化,增加抗炎M2亚型小胶质细胞的数量,同时在脑组织内上调抗炎因子TGF-β和IL-10的表达,这种免疫调节作用也参与了MSC-EVs对AD神经元的保护作用。在AD转基因小鼠的体内证实,MSC-EVs治疗能显著延缓AD认知功能缺陷的病程发展。因此,这种认知保护作用对减缓AD患者在病程中的生活质量下降将具有重要意义。不同的培养环境对干细胞表型及SC-EVs成分具有明显影响。
最新研究发现在缺氧条件培养下MSCs分泌的MSC-EVs中miR-21的表达水平明显升高。通过与miR-21-MSC-EVs在AD转基因小鼠体内的治疗效果进行对比,该研究发现miR-21+MSC-EVs能够调节促炎-抗炎因子平衡,并促进神经元突触与轴突生长,而正常条件培养所分泌的miR-21-MSC-EVs不具有明显的这些现象。由于低氧的微环境也存在于AD的病理过程中,这项结果说明miR-21可能参与了MSCs对AD的治疗作用,且通过MSC-EVs运载该结论对未来更有效地开展AD治疗具有重要启发。在Aβ蛋白的AD模型中,MSC-EVs具有降解Aβ蛋白并保护神经的治疗作用,在动物体内也具有确切的认知功能保护疗效。
AD作为一种非损伤性的中枢神经系统病变,MSC-EVs的非移植性及安全性是其在未来实际AD临床治疗中的巨大优势。但对于目前的研究结果而言,MSC-EVs未能从实际老化引起的AD模型中取得疗效成果,仅从Aβ蛋白的角度证实的MSC-EVs疗效可能需要在未来的临床试验中作进一步的验证。
2.3SC-EVs对神经胶质瘤的治疗
神经胶质瘤是一种常见的中枢神经系统恶性肿瘤。尽管手术切除是目前胶质瘤的主要治疗方法,但对于恶性肿瘤常见的浸润性病灶,手术无法达到完全切除,需结合术后化疗等进行综合治疗。而胶质瘤的化疗也存在着血脑屏障阻隔、肿瘤细胞对药物不敏感的难题,因此急需寻找一种新的高效治疗方式。近年来,MSCs被发现对多种恶性肿瘤的生长具有明显的抑制作用。大量研究发现,MSCs移植后对脑内胶质瘤部位具有趋向性,在达到肿瘤部位后MSCs能通过旁分泌MSC-EVs等因子来调节肿瘤细胞微环境,从而抑制肿瘤细胞增殖并诱导细胞凋亡。
由于MSC-EVs比MSCs在治疗上更安全可行,因此研究MSC-EVs对胶质瘤的治疗效果成为探索一种新的治疗方式的途径。在对其他恶性肿瘤的治疗研究中发现,MSC-EVs中含有的miRNA是抑制肿瘤生长的关键作用分子。有研究证实,miR-124可以通过抑制肿瘤细胞的STAT3通路,减少肿瘤细胞的免疫逃逸,起到抗肿瘤的治疗效果。
在神经系统最常见的恶性肿瘤胶质瘤的治疗中,近期研究发现miR-584+MSC-EVs能使体外胶质瘤细胞中与肿瘤发生相关的MMP-2蛋白的表达下降60%。并且miR-584还能激活下游caspase-3促进肿瘤细胞凋亡。在胶质瘤小鼠模型体内也发现,miR-584+MSC-EVs明显抑制体内胶质瘤的生长,具有显著的抗肿瘤疗效。在另一项研究中发现,通过MSC-EVs载运,MSCs所分泌的miR-124a能够大量进入颅内病灶并被胶质瘤细胞所吸收。miR-124a通过抑制肿瘤细胞的FOXA2的表达来抑制细胞增殖及肿瘤生长。对胶质瘤小鼠模型的在体治疗发现,miR-124a+MSC-EVs可以显著延长肿瘤小鼠的平均生存时间,具有巨大的胶质瘤治疗价值。
ESCs是一种多能干细胞,具有分化为多种组织的潜力。但与MSCs这类成体干细胞不同的是,ESCs因具有多能潜力,会在体内移植后形成畸胎瘤样的良性肿瘤。这是判断ESCs在体内移植后是否存活的标准,但也限制了ESCs在体内的治疗使用。由于ESCs同时具有更强的自我更新和增殖能力,能够分泌大量性质稳定的ESC-EVs,而ESC-EVs不会在体内移植后形成畸胎瘤,因而是ESCs的一种安全有效的替代方案。
近期有研究通过将ESCs与癌细胞共培养,发现ESCs通过分泌ESC-EVs传递mRNA,对乳腺癌细胞与结直肠癌细胞进行基因再编辑,从而抑制癌细胞增殖。证实ESC-EVs对恶性肿瘤具有抑制生长的作用。在此基础上,作者汪泱团队的最新研究发现,ESC-EVs能够抑制体外胶质母细胞瘤细胞生长,对动物体内的胶质瘤生长也具有显著抑制作用。
研究团队将靶向胶质母细胞瘤细胞的分子标志cRGyDK插入ESC-EVs质膜,并使ESC-EVs携带紫杉醇并被胶质瘤细胞吸收,进一步增强了ESC-EVs对肿瘤细胞的靶向性与杀伤性。改造后ESC-EVs具有更强的肿瘤疗效。此外,由于ESC-EVs容易被细胞吸收,并在改造后具有更强肿瘤趋向性,ESC-EVs携带的紫杉醇能以较低剂量达到单用紫杉醇的同等化疗效果,减少了化疗药物的剂量依赖副作用发生。该研究在动物体内体外完整证实了ESC-EVs对胶质母细胞瘤的治疗作用,同时也证明了ESC-EVs的抗肿瘤作用具有强大可塑性,具有巨大的研究前景。