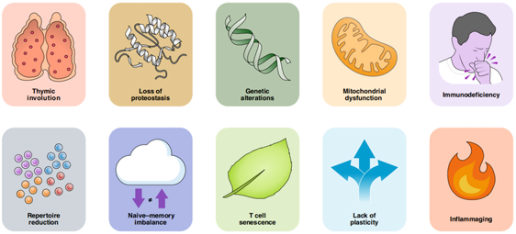
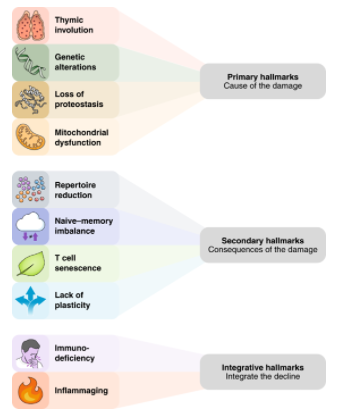
1、胸腺退化
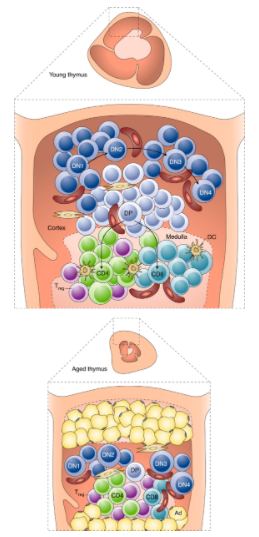
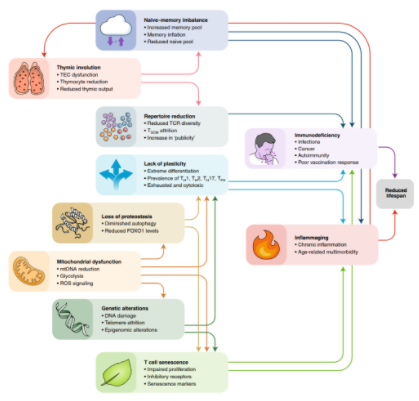
免疫系统发生衰老,最有据可查的变化就是胸腺的退化,胸腺是负责产生 T 细胞的主要器官。人类胸腺的退化开始于童年,退化巅峰于青春期。与年龄相关的胸腺退化表现为组织结构的破坏、胸腺质量的减少以及胸腺细胞数量的下降。这导致到达外周血里的初始T细胞减少,记忆T细胞的代偿性克隆性扩增,外周T细胞库多样性减少,影响了对病原体的检测。
性类固醇水平的增加和生长因子(如生长激素、IGF-1和KGF)的产生减少,可促进胸腺退化。因此,通过手术或化学阉割导致的性类固醇减少,可诱导胸腺功能的恢复。相反,压力、感染、肥胖、怀孕和抗肿瘤治疗会加速胸腺退化。
在化疗、电离辐照和感染引起损伤后,老年人无法恢复免疫功能,从而增加了疾病的发病率和死亡率。因此,制定增强胸腺功能和促进免疫重建的策略至关重要。通过额外施用生长激素/瘦素/生长激素释放肽(ghrelin)等可防止压力介导的胸腺萎缩。注射Ghrelin可显著增大年老小鼠的胸腺、提升细胞数量和T细胞输出。Ghrelin输注可以使老年小鼠胸腺大小、细胞数量和T细胞输出显著增加。另外,IL-22也可能促进小鼠胸腺再生。值得注意的是,经常锻炼的人初始T细胞和新迁出胸腺细胞(RTE)的频率增加,以及IL-7的循环水平升高。除了激素和生长因子外,胸腺移植和过继细胞治疗也可能刺激小鼠胸腺发育。
值得注意的是,胸腺萎缩和胸腺输出减少的机制在很大程度上仍然是难以捉摸的。阐明这些关键途径可能为预防免疫衰老和延缓炎症反应的新策略铺平道路。一句话:胸腺强则免疫强,胸腺弱则免疫弱。
2、线粒体功能障碍
线粒体功能障碍大多发生在衰老的组织和细胞类型中(当然也包括T细胞)。比起年轻个体,老年个体的T细胞中含有更丰富的线粒体蛋白,但它们被氧化磷酸化而受损,这表明细胞中累积着大量功能失调的线粒体。究其原因可能是线粒体的自噬的无效循环造成的。
线粒体不仅对生物能量学和细胞代谢(包括维持NAD/NADH比值)至关重要。线粒体还是产生、传递和响应活性氧(ROS)或钙峰的信号中枢。而在老化的T细胞中,这些信号通路失调。T细胞线粒体功能障碍通过多种分子机制(包括炎症代谢物的积累、表观遗传改变、转录后蛋白修饰),导致促炎症表型的获得。在T细胞中,线粒体衰退与衰老表型的获得有关。因此,低剂量鱼藤酮可以抑制线粒体中呼吸链复合物I,加速CD4+T细胞的免疫衰老。此外,敲除T细胞线粒体转录因子A(英文缩写:TFAM),可导致线粒体衰竭。
目前,二甲双胍正在开展延缓衰老的临床试验。使用二甲双胍,可增强小鼠CD8+T细胞中的TFAM表达和线粒体功能,有助于解决结核分枝杆菌的感染。线粒体功能恢复增强策略已被用于恢复耗尽的TIL。GDF15是一种线粒体应激或功能障碍产生的线粒体因子。GDF15可以维持Treg细胞的免疫抑制功能并减缓炎症。干预内分泌因素(包括GDF15)可能为改善老年人免疫提供一种新的策略。
一句话:恢复线粒体功能可能有助于T细胞功能紊乱的恢复。
3、遗传和表观遗传的改变
随着年龄增长,T细胞也会发生遗传和表观遗传改变。染色体变化在老化的T细胞中积累,T细胞白血病发病率的增加,在这方面日本人比欧洲人更易发生。T细胞基因组不稳定可能是由线粒体应激,ROS的过度产生和端粒磨损以及修复酶的活性降低导致的。所有主要的T细胞亚群,都表现出与年龄相关的端粒长度减少,这与人类中受到慢性感染关系很大,尤其是巨细胞病毒(CMV)。特发性肺纤维化(IPF)是人类短端粒综合征最常见的表现。接受肺移植的IPF患者T细胞对CMV的免疫力受损,证实端粒变短会损害T细胞功能。因此,端粒酶突变可能会发展为T细胞免疫缺陷。越接近健康的百岁老人,T细胞端粒越长、端粒酶活性越高和细胞增殖能力越强。表观遗传改变可能一直伴随着T细胞衰老。其影响DNA甲基化以及组蛋白编码及其多重翻译后修饰,这在一定程度上解释了T细胞的老化过程。在白细胞端粒长度较短的个体中,DNA甲基化年龄特别高,与低水平的记忆CD8+T细胞和高水平的初始CD8+T细胞相关。据报道,DNA甲基组与年龄相关的表观遗传变化在30岁末或40岁初首次激增,时间和女性相似;第二次高峰更早(5-6年后),男性更强,这与男性预期寿命的降低有关。一句话:不论是遗传还是表观遗传,均可能造成 T 细胞衰老,进而影响整体健康状况,甚至寿命。
4、蛋白稳定性丧失
对于错误折叠或老化蛋白质的降解,机体是分两种途径介导的,分别是蛋白水解酶和自噬。这两个任何一个出现缺陷都会导致T细胞老化。自噬是机体内大蛋白聚集体和细胞器循环利用的唯一机制,自噬体通过与溶酶体融合以大量降解管腔内容物。随着年龄的增长,自噬功能趋于不足,这些因素可能包括:伴随代谢综合征的营养物质(葡萄糖和游离脂肪酸)和生长因子(胰岛素和IGF1)的增加,过度激活抑制自噬的mTORC1通路;
由于烟酰胺二核苷酸(NAD)的年龄相关性下降,营养传感器(如Sirtuin-1)的激活水平降低;
降低了促自噬蛋白的催眠依赖性翻译所需的亚精胺水平;
降低了自噬转染因子级联的激活,如FOXO1-TFEB轴。
从理论上讲,自噬可以通过减少热量摄入(通过热量限制或间歇性禁食)和提供NAD前体(如烟酰胺、精胺)膳食补充剂来重建,从而增加健康寿命和寿命。自噬的年龄依赖性下降也会影响T细胞。记忆T细胞和Treg细胞比效应T细胞更依赖氧化磷酸化,特别容易受到自噬抑制的影响。自噬的药理学抑制会干扰人类记忆CD8+ T细胞的组织驻留重编程。自噬具有至少部分由Treg细胞介导的抗炎作用。雷帕霉素(mTORC1抑制剂)的短期给药可提高小鼠CD8+ T记忆细胞对病毒感染的免疫反应,并提高老年人对流感病毒疫苗的免疫反应。在属于长寿家庭的个体中,与对照组相比,CD4+ T细胞显示出改善激活诱导的自噬活性。总之,自噬是对抗T细胞衰老的有效靶点,诱导自噬可以减少炎症同时提高免疫反应的灵敏度。一句话:自噬作用恢复或增强有效延缓衰老,控制热量摄入和间歇性饮食是个好主意。另外,NAD前体、精胺、雷帕霉素也值得研究。